ARTÍCULO ORIGINAL
Zumbido urbano: implicancia de polinizadores silvestres en la carga polínica de Solanum lycopersicum en Rosario
Urban buzz: role of wild pollinators in pollen load on Solanum lycopersicum in Rosario
Zumbido urbano: implicação dos polinizadores silvestres na carga poliníca de Solanum lycopersicum em Rosario
Mazzei,
Mariana Paola1,2![]() ; Galetto,
Leonardo3
; Galetto,
Leonardo3![]() ;
Vesprini, José Luis1,2
;
Vesprini, José Luis1,2![]()
1Cátedra de
Ecología, Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Campo
Experimental J. F. Villarino, 2125 Zavalla, Santa Fe, Argentina.
2 Instituto de Investigaciones en Ciencias Agrarias, IICAR-CONICET,
Campo Experimental J. F. Villarino, 2125 Zavalla, Santa Fe, Argentina.
3 Instituto Multidisciplinario de Biología Vegetal,
IMBIV-CONICET-Universidad Nacional de Córdoba, Vélez Sarsfield 1666, 5016
Córdoba, Argentina.
mariana.p.mazzei@gmail.com
DOI: https://doi.org/10.35305/agro46.e054
Recibido: Julio 2025 Aceptado: Noviembre 2025
Resumen
La fauna de polinizadores silvestres en huertas periurbanas de Rosario (Santa Fe, Argentina) permanece prácticamente desconocida, a pesar de que numerosos cultivos, como Solanum lycopersicum var. platense (tomate), mejoran su producción con estos insectos. Aunque el tomate produce frutos por autopolinización, la polinización mediada por insectos mejora notablemente la producción de frutos. Este estudio tuvo dos objetivos principales: i) elaborar un inventario de polinizadores silvestres en huertas periurbanas de tomate, y ii) cuantificar el depósito de polen en los estigmas provocado por la visita de insectos. Se realizaron observaciones en 60 inflorescencias de tomate en 4 huertas, registrando la abundancia, frecuencia y comportamiento de los visitantes florales. Se comparó la cantidad de polen depositado sobre estigmas de flores expuestas a los visitantes con la cantidad de polen depositado sobre estigmas de flores excluidas de visitantes (n=96). Se identificaron nueve morfoespecies de las cuales cinco correspondieron a abejas capaces de realizar polinización por zumbido (buzz‑pollination), destacándose la familia Halictidae con una abundancia de 107 visitas. Las abejas que recolectan polen por zumbido representaron la mayoría de las visitas. Los estigmas expuestos mostraron más del doble de granos de polen que los estigmas de flores excluidas. Las huertas periurbanas pueden estar actuando como sitios de alimentación para polinizadores nativos, lo cual trae aparejado aumentos en el proceso de polinización.
Palabras claves: abejas; polen; tomate
Summary
The wild pollinator fauna in peri-urban vegetable gardens of Rosario (Santa Fe, Argentina) remains largely unknown, despite the fact that numerous crops, such as Solanum lycopersicum var. platense (tomato), benefit from the activity of these insects. Although tomatoes can produce fruits through self-pollination, insect-mediated pollination significantly enhances fruit production. This study had two main objectives: (i) to compile an inventory of wild pollinators in peri-urban tomato gardens, and (ii) to quantify pollen deposition on stigmas caused by insect visits. Observations were conducted on 600 tomato inflorescences in four gardens, recording the abundance, frequency, and behavior of floral visitors. The amount of pollen deposition on stigmas of flowers exposed to visitors was compared with that on stigmas of flowers excluded from visitors (n = 96). Nine morphospecies were identified, five of which corresponded to bees capable of performing buzz pollination, with the family Halictidae standing out for its abundance (107 visits). Bees that collect pollen through buzzing accounted for the majority of visits. The exposed stigmas showed more than twice as many pollen grains as the stigmas of excluded flowers. Peri-urban gardens may function as foraging sites for native pollinators, contributing to an increase in pollination.
Keywords: buzz-pollinating bees; pollen; tomato
Resumo
A fauna de polinizadores silvestres em hortas periurbanas de Rosario (Santa Fe, Argentina) permanece praticamente desconhecida, apesar de diversas culturas, como Solanum lycopersicum var. platense (tomate), melhorarem sua produção com esses insetos. Embora o tomate possa produzir frutos por autopolinização, a polinização mediada por insetos aumenta significativamente a produção de frutos. Este estudo teve dois objetivos principais: (i) elaborar um inventário de polinizadores silvestres em hortas periurbanas de tomate e (ii) quantificar o depósito de pólen nos estigmas causada pelas visitas de insetos. Foram realizadas observações em 60 inflorescências de tomate em quatro hortas, registrando a abundância, frequência e comportamento dos visitantes florais. Foi comparada a quantidade de pólen depositado nos estigmas das flores expostas aos visitantes com a quantidade de pólen depositado nos estigmas das flores excluídas dos visitantes (n=96). Nove morfoespécies foram identificadas, cinco das quais corresponderam a abelhas capazes de realizar polinização por zumbido (buzz‑pollination), destacando-se a família Halictidae pela sua abundância (107 visitas). As abelhas que coletam pólen por zumbido representaram a maioria das visitas. Os estigmas expostos apresentaram mais do que o dobro de grãos de pólen em comparação com os estigmas das flores excluídas. As hortas periurbanas podem funcionar como locais de alimentação para polinizadores nativos, contribuindo para o aumento do processo de polinização.
Palavras-chave: abelhas; pólen; tomate
Introducción
La polinización por vibración o zumbido (“Buzz-pollination”) o sonicación floral es el proceso de liberación de polen de las anteras de la flor por medio de vibraciones torácicas por parte de los visitantes florales ( Rosi‐Denadai et al., 2020). La mayoría de las especies del género Solanum L. poseen flores con anteras poricidas que liberan el polen a través de poros apicales, extraído mediante la polinización por zumbido (Carrizo Garcia et al., 2008). Esta acción es realizada por ciertas abejas que recolectan el polen como recompensa y lo utilizan para la alimentación de sus larvas (De Luca y Vallejo-Marín, 2013; Buchmann, 1985). Las flores de tomate Solanum lycopersicum L. son visitadas por muchos insectos como coleópteros, dípteros y abejas, aunque la mayoría son abejas que vibran sobre las flores para obtener los granos de polen (e.g., Herwig, 2005; Vinícius-Silva et al., 2017). Cuando este tipo de abejas se posa sobre la flor, curva el lado ventral de su cuerpo sobre las anteras, mientras se aferra con las mandíbulas a la base de la flor; luego, desacoplan el mecanismo de vuelo dejando de batir las alas y contrayendo rápidamente los músculos torácicos. Las vibraciones resultantes se transmiten a las anteras a través de la cabeza, las mandíbulas y el lado ventral del abdomen, causando que los granos de polen ganen energía y sean expulsados a través de los poros apicales de cada antera (De Luca y Vallejo-Marín, 2013). El polen expulsado se deposita en el cuerpo del polinizador donde luego puede ser preparado y recogido para transportarlo de regreso al nido o depositado sobre el estigma de otras flores de tomate contribuyendo a la polinización cruzada (De Luca y Vallejo-Marín, 2013).
Las flores en antesis de S. lycopersicum tienen una longevidad floral de 7 días y pueden alcanzan hasta un 98 % de autopolinización (Aldana et al., 2007). Este tipo de polinización, principalmente por acción del viento que mueve las flores y la gravedad, produce frutos de menor calidad que la polinización cruzada (Aldana et al., 2007; Gabri Martín y Cuesta, 2013; Cooley y Vallejo-Marín, 2021; Corti et al., 2024). Se ha demostrado que la polinización cruzada, entre distintas flores mediada por abejas que recolectan polen por zumbido, mejora la producción de frutos y semillas (Cooley y Vallejo-Marín, 2021). El tamaño del fruto de tomate y su calidad dependen directamente de la cantidad de granos de polen que se depositan sobre el estigma; así, a menor cantidad de polen, se producen pocas semillas, frutos más pequeños, livianos y poco uniformes (Morandin et al., 2001; de Melo et al., 2013; Wongsa et al., 2023). Se observaron además aumentos en los parámetros productivos del cultivo cuando se incrementan las visitas de los polinizadores a las flores (Depra et al., 2014; Arias Restrepo et al., 2015; Cerna Chávez et al., 2015; Corti et al., 2024), por lo que se ha clasificado a este cultivo como moderadamente dependiente a la polinización biótica mediada por insectos. Aunque muchos trabajos en huertas de tomate relacionan la cantidad de visitas de las abejas a las flores con la producción de frutos, pocos han evaluado la cantidad de polen depositado en los estigmas como medida del éxito reproductivo (Morandin et al., 2001; de Melo et al., 2013; Corti et al., 2024). La limitación por polen puede deberse tanto a la cantidad (escaso número de granos capaces de fertilizar todos los óvulos) como a la calidad (granos de polen que, debido a factores genéticos, compatibilidad o interacciones con otros granos de polen, no logran producir semillas viables); ambas ejercen efectos negativos sobre la fecundidad de los óvulos, la formación de semillas y el desarrollo de los frutos (Aizen y Harder, 2007; Lee, 1988). Husband y Schemske (1996) reportaron que, en comparación con la polinización cruzada, la autopolinización redujo la producción de semillas en un promedio del 20 % en 62 especies o poblaciones autocompatibles, alcanzando una reducción máxima del 87 %. Un mayor número de granos de polen en el estigma no solo aumenta las probabilidades de fertilizar todos los óvulos, sino que también incrementa la probabilidad de recibir polen de mayor calidad. En diversos cultivos autocompatibles se ha observado una relación positiva entre la cantidad de polen y el desarrollo de los frutos al comparar tratamientos de exclusión de flores a polinizadores (autopolinización) y exposición de flores a polinizadores (autopolinziación+polinización cruzada) (Corti et al., 2024 (tomate); Mazzei et al., 2021 (colza); Santone et al., 2022 (soja)).
El periurbano de Rosario es una de las áreas hortícolas más grandes de Argentina abasteciendo de hortalizas frescas a la población de la ciudad de Rosario, con la ventaja de la cercanía entre el productor y el consumidor; en el caso de tomate, su producción es al aire libre y se vende para el consumo fresco (Municipalidad de Rosario, 2020, 2022; Mondino et al., 2021). Es importante llevar a cabo investigaciones sobre el ensamble de polinizadores presente en estos sitios, con el fin de conocer la biodiversidad local y desarrollar en un futuro estrategias que favorezcan el servicio ecosistémico de la polinización biótica. Esto resulta especialmente relevante ante la creciente demanda de productos hortícolas en las ciudades y la necesidad de promover prácticas más sostenibles en las áreas periurbanas (Bracalenti et al., 2019).
Este trabajo tuvo como objetivos I).Generar un listado de potenciales polinizadores silvestres del tomate redondo (Solanum lycopersicum var. platense ) en huertas del periurbano de Rosario; y II). Evaluar el depósito de polen en el estigma de flores expuestas a los visitantes florales (principalmente las abejas que recolectan polen por zumbido) en comparación a flores excluidas. Esto permitirá estimar la influencia de los polinizadores silvestres en la mejora de la polinización del cultivo.
Materiales y métodos
Se trabajó en cuatro huertas del periurbano del departamento Rosario, dos en la localidad de Zavalla (33°01´52.77´´S 60°53´48.88´´O y 33°01´48.93´´S, 60°53´04.19´´O), una de las cuales está inmersa en un sistema de producción tradicional y otra dentro de un módulo agroecológico. Las dos restantes fueron: una huerta orgánica ubicada en la localidad de Funes (32°53´32.12´´S, 60°49´52.20´´O) y otra de transición agroecológica ubicada en Pueblo Esther (33°04´04.41´´S, 60°34´30.18´´O). En cada huerta se trasplantaron 72 plantines de S. lycopersicum var. platense (tomate redondo) generados en la Facultad de Ciencias Agrarias. Los plantines se llevaron a campo cuando alcanzaron una altura aproximada de 10 cm (tres semanas).
Visitantes florales y potenciales polinizadores
Durante el período de floración, se observaron 30 inflorescencias de tomate de plantas distintas por día en 5 días por huerta (n total=600). Cada inflorescencia se observó por diez minutos. Los días y las horas de registro dentro del rango de las 8:00 a 14:30 h fueron seleccionados según las condiciones climáticas óptimas para la observación de polinizadores (velocidad del viento menor a 20 km/h y cielo despejado). De cada observación se registró la identidad de los visitantes florales y el número de flores abiertas por inflorescencia. Se fotografiaron y capturaron los ejemplares para su posterior determinación taxonómica hasta el mínimo nivel taxonómico posible. De cada morfoespecie de visitante se observó el máximo de visitas por inflorescencia durante 10 minutos y se calculó la abundancia como: número total de visitas observadas por tiempo total de observación, y la frecuencia relativa de visitas por flor como:

Además se examinó el comportamiento de forrajeo de los visitantes sobre las flores siguiendo a De Luca y Vallejo-Marín (2013) y se determinó si el visitante floral realizó una recolección de polen por zumbido . A partir, de la frecuencia de visita y el comportamiento observado en las flores, diferenciamos entre polinizadores potenciales y visitantes florales. Se consideraron polinizadores potenciales aquellas especies que, además de presentar altas frecuencias de visita (mayor a 10 %), realizaron contacto directo con las estructuras reproductivas de la flor (estigma y/o anteras) y fueron capaces de efectuar recolección de polen por zumbido. En cambio, se clasificaron solo como visitantes florales aquellas especies que, aunque presentes en las inflorescencias, mostraron frecuencias de visita bajas (menor a 10 %) y/o no realizaron contacto efectivo con estigmas o anteras, y/o no recolectaron polen por zumbido, por lo que su influencia potencial en la polinización se consideró irrelevante.
Depósito de polen en los estigmas
En cada huerta se marcaron al azar 24 inflorescencias, 12 de ellas se cubrieron con bolsas de voile (30 x 15 cm) excluyendo a los visitantes florales (EVF), y los otros 12 se marcaron en el pedúnculo con un hilo de color, sin excluir a los visitantes florales (SEVF). Al final de la antesis se colectaron las inflorescencias de cada tratamiento y todos los gineceos se conservaron en FAA (Formaldehído, Alcohol, Ácido acético; 10:50:5, completando con 35 ml de agua). Se eligió al azar un estigma por inflorescencia de cada tratamiento de polinización (n=12 por tratamiento por huerta, n=96 total). Luego, se sometió a la técnica de Squash (NaOH 8N por 10 minutos a 60°C) y a una tinción con safranina (DAFNI et al., 1992). Se observaron los preparados con microscopio estereoscópico Leica Diastar, (objetivo10x) y se tomaron fotografías. Para el conteo comparativo entre tratamientos de granos de polen se eligió una foto central de cada preparado de estigma en la cual toda la superficie correspondía a tejido estigmático. Mediante la función: “contador de células” del software ImageJ, se realizó el conteo visual de los granos de polen presentes en una superficie estigmática de 0.44 mm2. Esta superficie se eligió para uniformizar el área de muestreo dentro del estigma ya que la técnica de squash genera distintas formas y superficies en el preparado. El conteo en el área constituyó la variable dependiente número de granos de polen depositados.
Análisis de datos
Se evaluó el efecto de los visitantes florales sobre el número de granos de polen depositados por estigma utilizando Modelos Lineales Generalizados Mixtos. Se incluyó planta seleccionada dentro de cada huerta anidada a la huerta como efecto aleatorio y el tratamiento (EVF o SEVF) se incorporó como efecto fijo. Luego de comparar las distribuciones de errores poisson y binomial negativa mediante la función fitdistr del paquete Fitdistrplus (Delignette-Muller y Dutang, 2015), evaluar los supuestos de ambos modelos ajustados con las funciones glmer y glmer.bn respectivamente del paquete lme4 (Bates et al., 2015) y la sobredispersión mediante el paquete DHARMa (Hartig, 2021) se seleccionó la distribución de errores binomial negativa para el modelo final. La media predicha por tratamiento y el intervalo de confianza del 95 % se calcularon con la función ggeffect del paquete ggeffects (Lüdecke, 2018). Todos los análisis se realizaron en el software estadístico R Project versión 4.4.2 (R Core Team, 2024) y los gráficos mediante el paquete ggplot2 (Wickham, 2016).
Resultados
Visitantes florales y polinizadores
Se observaron 167 visitantes florales pertenecientes a nueve morfoespecies de las Familias Apidae, Halictidae y Vespidae del Orden Hymenoptera y Syrphidae del Orden Diptera (Tabla 1). Cinco de las morfoespecies fueron abejas que realizaron la recolección de polen por zumbido (Bombus pauloensis, B. morio, Halictidae indet. 1, Augochloropsis indet. 1 y Eucerini indet. 1) y representaron el 55,55 % de los visitantes con una frecuencia por flor de 95,89 %. A lo largo de 10 minutos de observación, las flores recibieron en promedio menos de una visita (~0,3 visitas) de estas abejas. Los visitantes florales más abundantes y frecuentes fueron dos morfoespecies de abejas de la familia Halictidae (Halictidae indet. 1 y Augochloropsis indet. 1) (Figura 1) sumando ambas una frecuencia por flor de 68,56 %. Las morfoespecies del género Bombus (B. pauloensis y B. morio) representaron una frecuencia por flor de 22,98 % (Tabla 1).

Figura 1. Polinizadores himenópteros más abundantes y frecuentes observados en flores de tomate; a: Halicitdae indet. 1 con una frecuencia de 35,88 % b: Augochloropsi indet. 1 con una frecuencia de 34,45 %.
Tabla 1. Visitantes florales observados en flores de tomate redondo. De cada morfoespecie se detalla la taxonomía, abundancia total observada, el máximo de visitas por inflorescencia durante 10 minutos de observación, la frecuencia relativa de visitas por flor, comportamiento de cada morfoespecie sobre la flor y clasificación como polinizador potencial (PP) o visitante floral (VF). Apis mellifera fue observada en las flores de tomate, aunque fuera de los periodos de 10 minutos de observación que se utilizaron para calcular la frecuencia de visitas (por ello no aparecen datos en la tabla).

Se consideraron potenciales polinizadores a: Halicitade indet. 1, Augochloropsis indet. 1, Eucerini indet. 1 y B. pauloensis (Tabla 1), ya que presentaron las mayores frecuencias de visita, realizaron contacto directo con estigmas y/o anteras y efectuaron recolección de polen por zumbido. En cambio, el resto de los insectos registrados (n = 5 morfoespecies) se consideraron únicamente visitantes florales (Tabla 1).
Depósito de polen
La variable número de granos de polen depositados por estigma varió entre los valores de 7 a 441 granos de polen. El factor tratamiento fue significativo en el modelo ajustado (F = 46.31, P < 0.001). Las inflorescencias con flores sin exclusión de visitantes florales (SEVF) presentaron una media de 185 (IC 95 % = [146.1–232.9]) granos de polen y las inflorescencias excluidas de los visitantes florales (EVF) una media de 70 (IC 95 % = [52.5–93.3]) (estimado del modelo= 0.97, DE= 0.15, z= 6.66, P < 0.001) (Figura 2; Figura 3).

Figura
2.
Granos de polen depositados sobre el estigma de tomate redondo. A. Foto del
preparado de una flor expuesta a los visitantes florales. B. Foto del preparado
de una flor excluida a los visitantes florales. Con flechas negras se marcan 2
granos de polen en cada preparado.
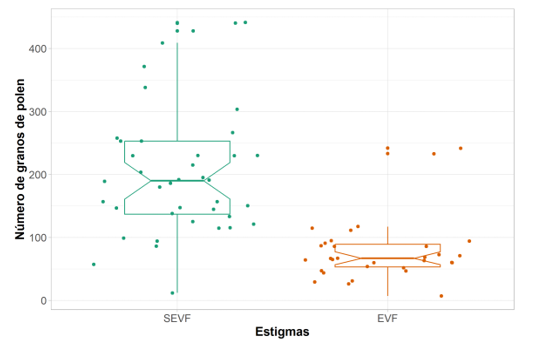
Figura 3. Número de granos de polen depositados sobre estigmas de flores de tomate redondo expuestas a los visitantes florales (SEVF) y excluidas de los visitantes florales (EVF). Los estigmas SEVF presentaron una mayor cantidad de granos de polen en su superficie que las flores asiladas (EVF), y esas diferencias fueron significativas (p< 0,001).
Discusión
Los resultados de este estudio permiten avanzar en el conocimiento de la fauna de visitantes florales y potenciales polinizadores silvestres en huertas periurbanas de Rosario. Se identificaron nueve morfoespecies de visitantes florales nativos, de las cuales cuatro fueron clasificadas como potenciales polinizadores del tomate redondo (Solanum lycopersicum var. platense), cumpliendo con el primer objetivo del trabajo. Asimismo, el depósito de polen en flores expuestas a visitantes florales aumentó significativamente en comparación con flores que solamente se autopolinizaron, lo que evidencia la contribución directa de las abejas silvestres en la mejora de polinización del cultivo y responde al segundo objetivo planteado. La comunidad de visitantes florales silvestres del tomate en huertas del departamento Rosario posee abejas con capacidad de recolección de polen por zumbido, siendo el grupo de visitantes más frecuentes sobre las flores. La tasa de visitas de las abejas registradas observada en periodos de 10 minutos indica que, si se toma en cuenta que las flores permanecen abiertas varios días, estas abejas son muy frecuentes y serían las responsables de la mayor cantidad de granos de polen depositados sobre los estigmas.
Si se considera además los resultados del experimento de exclusión de insectos visitantes, los potenciales polinizadores serían capaces de incrementar notablemente el depósito de polen en tomate redondo. De hecho, cuando las abejas accedieron a las flores los granos de polen depositados sobre los estigmas casi se triplicaron en comparación a las flores excluidas (autopolinización), coincidiendo esta tendencia con resultados previos en tomate (de Melo et al., 2013). Además, otros estudios en tomate han encontrado que cuanto mayor es la frecuencia de visitas y el depósito de polen, mayores son otros parámetros de rendimiento del cultivo como el peso, número de semillas y tamaño del fruto (e.g., Morandin et al., 2001; Corti et al., 2024). Sin embargo ambos estudios mencionados sólo trabajaron con categorías de niveles de polinización de abejas del género Bombus que fueron reconocidos por la decoloración necrótica del cono de anteras como resultado de la polinización por vibración realizada por abejorros (Morandin et al., 2001; Corti et al., 2024). Al aumentar el número de granos depositados sobre los estigmas se incrementa la cantidad de óvulos fecundados, lo que se relaciona positivamente con un número mayor de semillas (de Melo et al., 2013) mejorando la calidad del fruto (Vinícius-Silva et al., 2017). Las relaciones entre cantidad de polen, formación de semillas y tamaño de fruto tendrían una base genética importante ya que se ha demostrado que el número de semillas afecta la actividad del gen fw 2.2, que es responsable de la producción de estímulos para el crecimiento de las paredes del ovario y la formación de frutos (Tanksley, 2004). Futuros estudios en estas huertas podrían evaluar el impacto de este aumento en la cantidad de granos de polen con aumentos en el tamaño y peso de los frutos o incluso características organolépticas del fruto.
La calidad de la polinización por abejas depende de características de la vibración producida por el insecto como la amplitud, la duración y la frecuencia (Morandin et al., 2001; De Luca y Vallejo-Marín, 2013). En algunos estudios, las abejas de la familia Halictidae fueron los polinizadores más abundantes del cultivo de tomate (ver: Herwig, 2005; Arias Restrepo et al., 2015; Amala, 2017), no así en otras investigaciones (ver: Greenleaf y Kremen, 2006; de Melo et al., 2013; Depra et al., 2014; Vinícius-Silva et al., 2017) donde los géneros Exomalops, Anthophora y Bombus de la familia Apidae fueron los más representativos. Algunos autores señalaron que la recolección de polen por zumbido de abejas de tamaño pequeño, como los halíctidos traen aparejados menores rendimientos en tomate (número de semillas, tamaño y peso del fruto), en comparación con otras abejas de mayor tamaño (Cerna Chávez et al., 2015; Amala, 2017). Sin embargo, recientemente, se ha demostrado que la frecuencia de la vibración no está relacionada con el tamaño del cuerpo de los polinizadores, las abejas pequeñas pueden compensar su baja amplitud de zumbido durante la polinización aumentando la duración y frecuencia de sus visitas (Rosi‐Denadai et al., 2020). Por lo tanto, todas las abejas capaces de recolectar polen por zumbido registradas en este trabajo pueden tener un rol importante en la polinización de tomate.
A pesar de que en América del Sur Apis mellifera está ampliamente distribuida y domina las interacciones con cultivos dependientes de la polinización (Sáez et al., 2012), en este estudio su abundancia no fue importante ni tampoco en otras Solanáceas cultivadas en esta región (Alvarez et al., 2014). Esto puede explicarse porque las abejas de miel y otros polinizadores que no son capaces de zumbar para liberar el polen, como los sírfidos, se limitan a extraer pequeñas cantidades de polen en este tipo de flores cuando insertan sus piezas bucales en los poros de las anteras, por lo que prefieren otros recursos florales disponibles durante la estación de floración del tomate (De Luca y Vallejo-Marín, 2013; Alvarez et al., 2014). Además, A. mellifera puede desplazarse grandes distancias para visitar flores más atractivas y son fieles a las flores que visitan ya que propagan información olfativa dentro de la colmena y estas señales influencian el comportamiento de búsqueda de alimento del resto de las abejas (Fernández y Farina, 2001; Grüter et al., 2006). En cambio, las abejas que zumban sobre la flor pueden extraer rápidamente una gran proporción de polen de cada antera (Buchmann, 1985) y tener mayor impacto en la polinización de los cultivos que ofrecen polen como recompensa. La diferencia en la eficiencia para recolectar polen de plantas con anteras poricidas entre los dos tipos funcionales de abejas (aquellas que zumban y aquellas que no) puede explicar por qué las abejas que recolectan polen por zumbido predominan en el tomate, ya que pueden considerarse "funcionalmente especializadas" (De Luca y Vallejo-Marín, 2013).
Algunas de las biotecnologías disponibles para permitir una producción de frutos de alta calidad en tomate se basan en la cría y comercialización de especies de polinizadores que incrementen la polinización, como los abejorros Bombus impatiens o B. terrestris (Cerna Chávez et al., 2015). La comercialización de especies exóticas puede generar impactos negativos sobre la fauna nativa. Por ejemplo, B. terrestris, una especie generalista europea utilizada ampliamente en Chile como polinizadora de cultivos, se ha expandido y establecido en el sur de Argentina, provocando el decline de poblaciones del abejorro nativo B. dahlbomii (Geslin y Morales 2015). También se estima un crecimiento lineal de las poblaciones de B. terrestris en el sur de Argentina en los próximos años con posibles consecuencias negativas nuevas para abejorros nativos (Fontúrbel et al., 2021). En Argentina, actualmente se están utilizando colonias del abejorro nativo B. pauloensis en algunas producciones de tomate a gran escala (Cerna Chávez et al., 2015; Corti et al., 2024). Otra tecnología disponible es el agregado exógeno de reguladores de crecimiento a las flores (auxinas y giberelinas) (Corti et al., 2024). Además del costo asociado a esas hormonas, este método requiere una considerable inversión en mano de obra y tiempo, dado que implica realizar micro-pulverizaciones o inmersiones florales a lo largo de todo el período de floración (Martinez et al., 2016; Corti et al., 2024). Los frutos producidos por el estímulo de hormonas presentan menor cantidad de semillas, menor porcentaje de líquido y grados Brix (cantidad de sacarosa en solución) y mayor vacío en su contenido locular, lo que trasunta en un fruto menos apetecible para el consumo fresco (Gabri Martín y Cuesta, 2013; Cooley y Vallejo-Marín, 2021; Corti et al., 2024). La aplicación de giberelinas y auxinas inducen un desarrollo morfológico e histológico diferente del tejido resultante de los ovarios fecundados (Gabri Martín y Cuesta, 2013). Ambos métodos utilizados más frecuentemente (el agregado de colonias de abejorros o de reguladores de crecimiento) implican altos costos económicos a los pequeños productores y posibles costos ambientales, mientras que el manejo de la fauna de polinizadores locales, manteniendo sitios de nidificación, de refugio y alimentación para las abejas, puede mejorar la producción al mismo tiempo que se conserva la biodiversidad (Roulston y Goodell, 2011). La presencia de las abejas registradas en este trabajo podría traer beneficios económicos al tomate y a otras Solanáceas de interés comercial que también tienen anteras poricidas, que suelen cultivarse en la región (como todas las variedades de tomate, pimientos y berenjenas (Raw, 2000; Patricio et al., 2012; Jayasinghe et al., 2017) y a otros cultivos locales altamente dependientes de la polinización biótica (como calabazas, zapallitos, zucchinis, pepinos etc. (Andrews et al., 2007; de Melo et al., 2013; Koné et al., 2019; Malerbo-Souza et al., 2019; Mazzei et al., 2025). Futuros estudios deberían relacionar el depósito de polen con los rindes en tomate y construir redes de interacción cultivos locales-polinizadores.
Los espacios periurbanos son espacios valiosos para estudiar polinizadores nativos por la diversificación a nivel predial que genera un mosaico de ambientes diversos en el paisaje local y regional (Riat et al., 2015; Mazzei et al., 2024). Estudios que permitan cuantificar el estado actual de la biodiversidad de insectos benéficos como los polinizadores permitirán valorizar el servicio ecosistémico en estas áreas y desarrollar mejores estrategias de producción que utilicen menos insumos externos.
Agradecimientos
A los huerteros que nos permitieron realizar la investigación. A la Facultad de Ciencias Agrarias de la Universidad Nacional de Rosario y a los Ingenieros Mauricio P. Ortiz Mackinson, David Balaban y Rodolfo O. Grasso de la Cátedra de Horticultura. JV y MPM agradecen a SECyT (UNR) y LG agradece a la SECyT (UNC) por el financiamiento otorgado. Por último, agradecemos sinceramente la predisposición y los valiosos comentarios de los revisores anónimos, quienes contribuyeron de manera significativa a mejorar este manuscrito.
Declaración de Autoría (CRedit)
MPM: Investigación, Toma de datos, análisis formal, metodología, redacción, revisión y edición. LG: supervisión, revisión y edición. JLV: metodología, supervisión, revisión y edición
Referencias
AIZEN, M. A., HARDER, L. D. (2007) Expanding the limits of the pollen-limitation concept: effects of pollen quantity and quality. Ecology, 88(2), 271–281.
ALDANA, J., CURE, J.R., ALMANZA, M.T., VECIL, D., RODRÍGUEZ, D. (2007) Effect of Bombus atratus (Hymenoptera: Apidae) on tomato production (Lycopersicon esculentum Mill.) in greenhouse in Bogotá plateau, Colombia. Agronomía Colombiana, 25: 62–71.
ALVAREZ, L.J., LUCIA, M., RAMELLO, P.J., DEL PINO, M., ABRAHAMOVICH, A.H. (2014) Abejas asociadas a cultivos de berenjena (Solanum melongena L., Solanaceae) en invernadero del Cinturón Hortícola de La Plata, Buenos Aires, Argentina. Revista de la Facultad de Agronomía, La Plata, 113: 211–217.
AMALA, U. (2017) Role of native buzz pollinator bees in enhancing fruit and seed set in tomatoes under open field conditions. Journal of Entomology and Zoology Studies, 5: 1742–1744.
ANDREWS, E.S., THEIS, N., ADLER, L.S. (2007) Pollinator and herbivore attraction to Cucurbita floral volatiles. Journal of Chemical Ecology, 33: 1682–1691.
ARIAS RESTREPO, S.M., TORRES CARRERA, J.D., MOLINA RICO, L.J. (2015) Estructura de la comunidad polinizadora en un cultivo de tomate Lycopersicon esculentum Mill. (Solanaceae) y análisis multivariado de la calidad del fruto, Caldas – Colombia. Scientia et Technica, 20: 382.
BATES, D., MÄCHLER, M., BOLKER, B. & WALKER, S. (2015) Fitting Linear Mixed-Effects Models using lme4. Journal of Statistical Software, 67(1).
BRACALENTI, L., ZIMMERMANN, E., ONOCKO, A., MENDÍAZ, G., MOSCONI, P., VÁZQUEZ, J., CAZZOLI, I., PÉREZ, A., MACIEYKO, C., RITTERBUCH, B., VALIENTE, J., VAZQUEZ, F., TERRILE, R., CENTRO, C. (2019) Paisaje productivo en la ciudad de Rosario. Prestaciones posibles para la adaptación y la mitigación del cambio climático.
BUCHMANN, S.L. (1985) Bees use vibration to aid pollen collection from non-poricidal flowers. Journal of the Kansas Entomological Society, 58: 517–525.
CARRIZO GARCIA, C., MATESEVACH, M., BARBOZA, G. (2008) Features related to anther opening in Solanum species (Solanaceae). Botanical Journal of the Linnean Society, 158 (2): 344-354.
CERNA CHÁVEZ, E., LARA SÁNCHEZ, E.D., OCHOA FUENTES, Y., HERNÁNDEZ BAUTISTA, O., AGUIRRE URIBE, L.A., LANDEROS FLORES, J., FLORES CANALES, R. (2015) Comparación de cuatro especies entomófilas sobre parámetros agronómicos del fruto de tomate de invernadero. Revista Mexicana de Ciencias Agrícolas, 2241.
COOLEY, H., VALLEJO-MARÍN, M. (2021) Buzz-pollinated crops: a global review and meta-analysis of the effects of supplemental bee pollination in tomato. Journal of Economic Entomology, 114: 505–519.
CORTI, C., MEROI ARCERITO, F.R., FERNANDEZ DE LANDA, G., MAZZEI, M.P., FERNANDEZ DE LANDA, M., MAGGI, M., GALETTO, L. (2024) Tomato production under greenhouse conditions: bumblebees or hormones? Scientia Horticulturae, 326: 112747.
DAFNI, A. (1922) Pollination Ecology a practical approach. Toronto: Oxford University Press. 250p.
DELIGNETTE-MULLER, M. L., DUTANG, C. (2015) Fitdistrplus : An R package for fitting distributions. Journal of Statistical Software, 64(4), 1–34.
DEPRA, M.S., DELAQUA, G.G., FREITAS, L., GAGLIANONE, M.C. (2014) Pollination deficit in open-field tomato crops (Solanum lycopersicum L., Solanaceae) in Rio de Janeiro state, Southeast Brazil. Journal of Pollination Ecology, 12.
FERNÁNDEZ, P. C.; FARINA, W. M. (2001) Changes in food source profitability affect Nasonov gland exposure in honeybee foragers Apis mellifera L. Insectes Sociaux, 48(4), 366–371
GABRI MARTÍN, C.G., CUESTA, G. (2013) Efecto de distintas técnicas para el establecimiento de frutos en la producción de tomate bajo invernadero. Universidad Nacional de San Juan.
GESLIN, B., MORALES, C. L. (2015) New records reveal rapid geographic expansion of Bombus terrestris Linnaeus, 1758 (Hymenoptera: Apidae), an invasive species in Argentina. Check List, 11, 1620-1620.
GREENLEAF, S.S., KREMEN, C. (2006) Wild bee species increase tomato production and respond differently to surrounding land use in Northern California. Biological Conservation, 133: 81–87.
GRÜTER, C.; ACOSTA, L. E.; FARINA, W. M. (2006) Propagation of olfactory information within the honeybee hive. Behavioral Ecology and Sociobiology, 60(5), 707–715.
HARTIG, F. (2021) DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed Regression Models).
HERWIG, T. (2005) Pollinators of tomato, Solanum lycopersicum (Solanaceae), in Central Europe. Phyton, 45: 217–235.
HUSBAND, B. C., SCHEMSKE, D. W. (1996) Evolution of the magnitude and timing of inbreeding depression in plants. Evolution, 50(1), 54–70.
JAYASINGHE, U.J.M., R., S., SILVA, T.H., E., S., KARUNARATNE, W.A., P., I. (2017) Buzzing wild bee visits enhance seed set in eggplant, Solanum melongena. Psyche: A Journal of Entomology, 1–7.
KONÉ, K., TUO, Y., YAPO, M.L., TRAORÉ, D., SORO, F., KOUA, K.H. (2019) Diversity, abundance and activity of bees in zucchini (Cucurbita pepo L.) crops in northern Côte d’Ivoire. International Journal of Entomology Research, 4: 41–45.
DE LUCA, P.A., VALLEJO-MARÍN, M. (2013) What’s the ‘buzz’ about? The ecology and evolutionary significance of buzz-pollination. Current Opinion in Plant Biology, 16: 429–435.
FONTÚRBEL, F. E., MURÚA, M. M., VIELI, L. (2021) Invasion dynamics of the European bumblebee Bombus terrestris in the southern part of South America. Scientific Reports, 11(1), 15306.
LEE, T. D. (1988) Patterns of fruit and seed production. In J. LOVETT DOUST, L. LOVETT DOUST (Eds.), Plant reproductive ecology: Patterns and strategies (pp. 179–194). Oxford University Press.
LÜDECKE, D. (2018) ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. Journal of Open Source Software, 3(26), 772.
MALERBO-SOUZA, D.T., ANDRADE, M.O., MEDEIROS, N.M.G., SILVA, T.G., SIQUEIRA, R.A. (2019) Biodiversidade e comportamento forrageiro de abelhas na cultura da aboboreira italiana (Cucurbita pepo). Colloquium Agrariae, 15: 11–21.
MARTINEZ, S., GARBI, M., CARBONE, A., MORELLI, G., ARGERICH, C., PACHECO, R., PUCH, L. (2016) Aplicación de reguladores auxínicos: efecto sobre el cuajado de fruto en tomate para consumo fresco. Asociación Argentina de Horticultura, 35: 30–40.
MAZZEI, M.P., GALETTO, L., VESPRINI, J.L. (2025) The role of native and exotic bees in Cucurbita maxima var. zapallito pollination: the effects of pollination intensity and visits on commercial fruits. Apidologie, 56(1): 17.
MAZZEI, M.P., VESPRINI, J.L., BREEZE, T.D., IGLESIAS, M.D.R., GALETTO, L. (2024) Creating territorialized sustainability indices to evaluate pollination. Agricultural Systems, 219.
MAZZEI, M. P., VESPRINI, J. L., GALETTO, L. (2021) Seminatural habitats and their proximity to the crop enhances canola (Brassica napus) pollination and reproductive parameters in Argentina. Crop Science, 61(4), 2713–2721.
DE MELO, C., NETO, S., LIMA GOMES, F., GONÇALVES, B.B., BERGAMINI LIMA, L., DA SILVA E ELIAS, M.A., FRANCESCHINELLI, E.V. (2013) Native bees pollinate tomato flowers and increase fruit production. Journal of Pollination Ecology, 11: 41–45.
MONDINO, M.C., GRASSO, R., BALABAN, D., ORTIZ MACKINSON, M. (2021) Censo 2021 del cinturón hortícola de Rosario.
MORANDIN, L.A., LAVERTY, T.M., KEVAN, P.G. (2001) Effect of bumble bee (Hymenoptera: Apidae) pollination intensity on the quality of greenhouse tomatoes. Journal of Economic Entomology, 94: 172–179.
MUNICIPALIDAD DE ROSARIO (2020) Proyecto Cinturón Verde: informe anual de gestión y acción.
MUNICIPALIDAD DE ROSARIO (2022) Agricultura urbana Rosario. https://www.agriurbanarosario.com.ar/espacios-de-agricultura-urbana.php
PATRICIO, G.B., GRISOLIA, B.B., DESUOACUTE, I.C., MONTAGNANA, P.C., BROCANELLI, F.G., GOMIG, E.G., DE CAMPOS, M.J.O. (2012) The importance of bees for eggplant cultivations (Hymenoptera: Apidae, Andrenidae, Halictidae). Sociobiology, 59: 1037–1052.
R CORE TEAM (2024) R: a language and environment for statistical computing.
RAW, A. (2000) Foraging behaviour of wild bees at hot pepper flowers (Capsicum annuum) and its possible influence on cross pollination. Annals of Botany, 85: 487–492.
RIAT, P., LAMBARÉ, D.A., STAMPELLA, P.C., POCHETTINO, M.L. (2015) La diversificación del paisaje como aspecto clave de la sustentabilidad social de los sistemas agroecológicos campesinos. En: Memorias del V Congreso Latinoamericano de Agroecología, pp. 1–6.
ROSI-DENADAI, C.A., ARAÚJO, P.C.S., CAMPOS, L.A. DE O., COSME, L., GUEDES, R.N.C. (2020) Buzz pollination in Neotropical bees: genus dependent frequencies and lack of optimal frequency for pollen release. Insect Science, 27: 133–142.
ROULSTON, T. H.,GOODELL, K. (2011) The Role of Resources and Risks in Regulating Wild Bee Populations. Annual Review of Entomology, 56(1), 293–312.
SÁEZ, A., SABATINO, M., AIZEN, M.A. (2012) Interactive effects of large- and small-scale sources of feral honey-bees for sunflower in the Argentine Pampas. PLoS ONE, 7: e30968.
SANTONE, A., MAZZEI, M. P., VESPRINI, J., TORRES, C., AMARILLA, L. D., GALETTO, L. (2022) Pollination service and soybean yields. Acta Oecologica, 116, 103846
SARANDÓN, S.J. (2019) Potencialidades, desafíos y limitaciones de la investigación agroecológica como un nuevo paradigma en las ciencias agrarias. Revista FCA UNCUYO, 51: 383–394.
TANKSLEY, S.D. (2004) The genetic, developmental, and molecular bases of fruit size and shape variation in tomato. The Plant Cell, 16: 181–189.
VINÍCIUS-SILVA, R., PARMA, D. DE F., TOSTES, R.B., ARRUDA, V.M., WERNECK, M. DO V. (2017) Importance of bees in pollination of Solanum lycopersicum L. (Solanaceae) in open-field of the Southeast of Minas Gerais State, Brazil. Hoehnea, 44: 349–360.
WICKHAM, H. (2016) ggplot2: Elegant Graphics for Data Analysis. Springer, Verlag New York.
WONGSA, K., DUANGPHAKDEE, O., RATTANAWANNEE, A. (2023) Pollination efficacy of stingless bees, Tetragonula pagdeni Schwarz (Apidae: Meliponini), on greenhouse tomatoes (Solanum lycopersicum Linnaeus). PeerJ, 11: e15367.