ARTÍCULO ORIGINAL
Seed related traits in two cultivars of the megathermic forage species Panicum coloratum var. makarikariense in a temperate environment
Rasgos relacionados con la semilla en dos cultivares de la especie forrajera megatérmica Panicum coloratum var. Makarikariense en un ambiente templado
Características relacionadas a sementes em duas cultivares da espécie forrageira megatérmica Panicum coloratum var. makarikariense em um ambiente temperado
Moresco Lirusso, María Florencia1; Sacido, Mónica Benita2; Pratta, Guillermo Raúl 3; Felitti, Silvina Andrea1
1 Laboratorio de Biodiversidad Vegetal y Microbiana. Instituto de Investigaciones en Ciencias Agrarias de Rosario (IICAR - CONICET). Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Parque Experimental Villarino, S2125ZAA Zavalla, Santa Fe, Argentina.
2 Cátedra de Forrajes, Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Parque Experimental Villarino, S2125ZAA Zavalla, Santa Fe, Argentina.
3 Cátedra de Genética, Facultad de Ciências Agrárias UNR, IICAR – CONICET).
felitti@iicar-conicet.gob.ar
sfelitti@unr.edu.ar
DOI: https://doi.org/10.35305/agro38.e014
Recibido 26/07/2021 Aceptado 12/10/2021
Abstract
Panicum coloratum es una especie gramínea C4 nativa de Sudáfrica. En Argentina, se la valoriza por su producción forrajera de buena calidad en ambientes en los que los recursos son limitados. La cosecha de semillas esta dificultada por la madurez no uniforme de sus cariopsis dentro de la panoja y su rápida desarticulación. El objetivo de este trabajo fue evaluar el potencial de dos cultivares de Panicum coloratum para desarticulación de cariopses, germinación y caracteres relacionados a panoja con efectos sobre la producción de semillas en dos ciclos de cultivo (2017 y 2019). La desarticulación de cariopses fue evaluada semanalmente usando trampas para semillas. Los resultados mostraron diferencias significativas entre cultivares en ambos años. El pico de caída ocurrió en el mismo momento fenológico en ambos cultivares Respecto a la germinación, el comportamiento de ambos cultivares fue similar en ambos años y entre diferentes tratamientos aplicados. Para los caracteres relacionados a panoja, se encontró amplia variabilidad y correlaciones altas y positivas entre las variables evaluadas mediante un Análisis de Componentes Principales, tanto para ambos años como para el conjunto de datos. En todos los casos, las dos primeras Componentes Principales explicaron un alto porcentaje de la variabilidad total. La información generada en este trabajo permite concluir que cv. Kapivera presentó mayor retención y menor producción de semillas que cv. Bambatsi.
Palabras claves: dehiscencia; germinación; interacción GxA.
Resumen
Panicum coloratum es una especie gramínea C4 nativa de Sudáfrica. En Argentina, se la valoriza por su producción forrajera de buena calidad en ambientes en los que los recursos son limitados. La cosecha de semillas esta dificultada por la madurez no uniforme de sus cariopsis dentro de la panoja y su rápida desarticulación. El objetivo de este trabajo fue evaluar el potencial de dos cultivares de Panicum coloratum para desarticulación de cariopses, germinación y caracteres relacionados a panoja con efectos sobre la producción de semillas en dos ciclos de cultivo (2017 y 2019). La desarticulación de cariopses fue evaluada semanalmente usando trampas para semillas. Los resultados mostraron diferencias significativas entre cultivares en ambos años. El pico de caída ocurrió en el mismo momento fenológico en ambos cultivares Respecto a la germinación, el comportamiento de ambos cultivares fue similar en ambos años y entre diferentes tratamientos aplicados. Para los caracteres relacionados a panoja, se encontró amplia variabilidad y correlaciones altas y positivas entre las variables evaluadas mediante un Análisis de Componentes Principales, tanto para ambos años como para el conjunto de datos. En todos los casos, las dos primeras Componentes Principales explicaron un alto porcentaje de la variabilidad total. La información generada en este trabajo permite concluir que cv. Kapivera presentó mayor retención y menor producción de semillas que cv. Bambatsi.
Palavras-chave: dehiscencia; germinación; interacción GxA.
Resumo
Panicum coloratum é uma espécie gramínea do tipo C4 nativa da África do Sul. Na Argentina é valorizada por sua produção de forragem de boa qualidade em ambientes onde os recursos são limitados. A colheita das sementes é dificultada pela maturação não uniforme de suas cariopses dentro da panícula e sua rápida desarticulação. O objetivo deste trabalho foi avaliar o potencial de duas cultivares de Panicum coloratum para desarticulação de cariopses, germinação e caracteres relacionados à panícula com efeitos na produção de sementes em dois ciclos de cultivo (2017 e 2019). A desarticulação de cariopses foi avaliada semanalmente por meio de armadilhas de sementes. Os resultados mostraram diferenças significativas entre as cultivares nos dois anos. O pico de queda das sementes ocorreu no mesmo momento fenológico nas duas cultivares. Em relação à germinação, o comportamento das duas cultivares foi semelhante nos dois anos e entre os diferentes tratamentos aplicados. Foi encontrada ampla variabilidade e correlações altas e positivas para os caracteres relacionados à panícula, entre as variáveis avaliadas por meio da Análise de Componentes Principais, tanto para ambos os anos quanto para o conjunto de dados. Em todos os casos, os primeiros dois Componentes Principais explicaram uma alta porcentagem da variabilidade total. As informações geradas neste trabalho permitem concluir que a cv. Kapivera apresentou maior retenção e menor produção de sementes que a cv. Bambatsi.
Palabras-chave: deiscência; germinação; interação GxA
Introduction
The advancement of agriculture has led to a shift of livestock to marginal areas with less productive capacity due to climatic and edaphic limitations. These aspects restrict the choice of forage species to those that, due to their origin, have adaptation characteristics at the physiological, morphological and productive levels (quality and quantity of forage produced) (Pérez, 2005). Furthermore, the increase in summer temperatures, caused by global warming, has strengthened the interest in evaluating the productive and reproductive characteristics of new megathermic species. Panicum coloratum is a megathermic summer perennial, grass species, with C4-type photosynthetic metabolism, e which gives it greater CO2 capture efficiency. It has a habit of tussock growth, erect bushes with short rhizomes (rarely stoloniferous), of slow establishment, tolerates puddling, cold and slightly saline soils. It shows adequate performance in regions with drought cycles and periodic waterlogging (Fossati y Bruno, 1982).
It has a good production and presents high forage quality in December compared to other C4 grasses, which allows it to be conserved as deferred forage towards winter. The leaves and stems are highly palatable even in mature stages. It belongs to the Poaceae Family, Panicoideae Subfamily, Paniceae Tribe. Its African origin has allowed it to evolve in a tropical and subtropical environment, developing physiological and morphological adaptations that make this species more efficient in the use of light, temperature and water resources. It has been used as forage in various parts of the world. Two varieties are mainly recognized: P. coloratum var. coloratum and P. coloratum var. makarikariense, however the variety P. coloratum var. kabumlabula is also mentioned in literature. In Argentina the most widespread varieties of this species are P. coloratum L. var. coloratum and P. coloratum L. var. makarikariense Goossens. The megathermic forage species are good forage producers and, in turn, possess relatively good quality. The average annual dry matter production is around 8 tons per hectare and offers crude protein values of over 14% during the Argentinean summer season. In Argentina, out of 7.8 million hectares of forage pastures, 1.4 million corresponded to C4-type grasses, of which 130.500 were Panicum coloratum (INDEC, 2002).
Throughout history, the fruits have evolved through various mechanisms to mediate between their maturation and dispersal of their seeds. Dehiscence is the phenomenon by which the fruits reach maturity and open naturally to let out and spread the seeds. In grasses, the seed and fruit are welded to form the grain (or cariopse), so dehiscence implies the fall of the entire structure. In general, the harvested ears result in a large proportion of low weight and immature seeds. These seeds have low germination percentages, and the seedlings that manage to develop have less vigor than those from mature seeds (Tischler y Ocumpaugh, 2004).
From an anatomical point of view, seed dehiscence is a process conditioned by the appearance of two layers of abscission at the base of the spicule. The mechanisms that operate in this process are: 1) the disintegration of the middle lamella and part of the cell wall as a result of chemical changes, and 2) mechanical detachment due to weak tissue of the abscission layer (Burson et al., 1983). Differences between dehiscent and non-dehiscent plants were described in relation to the density of the abscission layer, the size of the cells of the sclerenchyma and parenchyma, the levels of hydrolytic enzymes and lignification (Patterson, 2001). Comparative anatomical and physiological studies in the Arabidopsis thaliana model plant revealed that dehiscence is very similar to the same process in Brassica (Spence et al., 1996).
The commercial seed production of tropical and subtropical perennial species has been really complex because they conserve wild characteristics that affect the harvest (Hacker y Loch, 1997), such as a caryopsis of caryopsis size that facilitates transportation by biotic and abiotic factors and an active dispersion mechanisms that cause the caryopsis to fall down (Hayes et al., 1989; Kennard et al., 2002; Cubero, 2003).
Since manipulation of dehiscence could improve crop yield, scientists have focused their attention on this process. Panicum coloratum var. makarikariense presents characteristics of determined flowering and uneven maturation within the panicle, which added to a high dehiscence of the seeds after their maturation; establish serious difficulties in harvesting quality seeds. Studies carried out by Dreher et al. (2008) on P. coloratum var. makarikariense showed that there are differences between different materials in terms of seed production per panicle, the number of empty seeds per panicle and the dynamics of seed fall.
The objective of this work was to study the dynamics of seed dehiscence, germination behavior and panicle attributes related to seed production and dispersion in two cultivars of Panicum coloratum var. makarikariense (cv. Kapivera and cv. Bambatsi) in the Pampeana Region of Argentina, with a temperated climate.
Materials and Methods
Plant material
Although P. coloratum var. makarikariense is a species originally collected from the Makarikari salt flats (now Makgadikgadi) on the African continent, the Bambatsi cultivar was launched in Australia in 1958 (Loch y Ferguson, 1999; Wilson, 1963). Kapivera is a cultivar developed by researchers from the National Institute of Agricultural Technology (INTA) Rafaela, Argentina. The most outstanding characteristic of this cultivar is its higher seed weight with respect to the original population. This results in more vigorous plants, with a higher dry weight, leaf length and total adventitious roots length. .Simultaneous to the development of Kapivera, different characters referring to panicle size were evaluated with the aim of reducing maturation asynchrony. During the selection process, no effects were found on the germination power of the seeds, however, it was highly influenced by the environmental influence (Maina, 2012).
During November 2015, plants of two cultivars of Panicum coloratum var. makarikariense (cv. Kapivera and cv. Bambatsi) were grown in plastic cups on a substrate composed of 50% soil and 50% vermiculite. After germination and two months of growth, the seedlings were transplanted into 2-liter bottles with soil and finally, in March 2016, 90 plants from each cultivar were established in the experimental field of the Faculty of Agricultural Sciences (National University of Rosario), located in Zavalla, Santa Fe, Argentina (33 ° 01 ′ S; 60 ° 53 ′ W). This locality presents a temperate climate with annual precipitations close to 990 millimeters, typical/vertic Argiudol soil made up of moderately well-drained fine clays and insufficiently provided with organic matter.Thelotconsists of two consecutive plots of identical dimensions with 1 meter separation between plants that were no previously chemical applied, neither during nor after transplant. Only two passes of a disk implement were made to promote soil movement and manual cleaning of the batch during the progress of the experiment.
Seed dehiscence
Seed drop was evaluated weekly for 70 days between March and May 2017 and 2019 by placing traps on 15 random inflorescences of each cultivar when 80% of them had reached 2/3 of flower opening. These traps consisted of a vertical iron structure covered by a lycra mesh to which two rings of equal diameter were attached at a distance of 30 centimeters between them, containing a seed collecting funnel at the base of the second ring (Figure 1). The inflorescences were incorporated into the trap by a slot made on the funnel, maintaining their natural conditions as far as possible. The collections were carried out until approximately 80% of the inflorescences had lost all of their seeds. Comparisons between cultivars for seed dehiscence were made using the X² non-parametric test (p = 0.05) (Di Rienzo et al., 2011).
Figure 1. Seed trap
used in this research.

Germination assay
For germination assay, 60 seeds of Panicum coloratum var. makarikariense cv. Kapivera and 150 seeds of Panicum coloratum var. makarikariense cv. Bambatsi were used. They were collected as described in the previous assay, during the austral autumn of 2017. The seeds were stored for no more than one year in paper envelopes in a cool and dry place until the moment of their use. The experiments were carried out approximately one year after having collected the seeds. Five different seed abrasion treatments and two growth conditions that presented variability in both luminosity and temperature were applied to scarify seeds (Table 1). The seed treatments performed were: one control without any abrasion on the integument (C), integuments remotion (WI), integuments scarified with sandpaper (SSP), seed coat subjected to the action of dry heat (DH), and seed coat subjected to the action of humid heat (HH). For WI treatment, a magnifying glass and scalpel was required to completely remove the seminal covering. The SSP treatment was carried out by placing the seeds inside a small jar internally surrounded with fine sandpaper, making circular movements for 5 minutes. Regarding the DH and HH treatments, the seeds were subjected to 60 ° C, in an oven and in water, respectively, for 5 minutes in each case. One growth conditions was with 12 hours of photoperiod and 20-25 °C, in a controlled growth chamber () and the other inside a greenhouse under natural conditions. Three replicates per sample were performed to increase the certainty of the results. After abrasion treatments, seeds were sown on filter paper moistened with distilled water inside glass Petri dishes and were placed under the different growth conditions. For the Kapivera cultivar, two seeds were used per replicate (six seeds for each treatment), for the Bambatsi cultivar, five seeds per repetition were used (fifteen seeds per treatment). The determination of germinated seeds was carried out daily for 10 days. Seeds were considered germinatedwhen the radicle was visible at approximately 1 millimeter. Comparisons between cultivars for each treatment and between treatments within each cultivar were performed using the χ² nonparametric test (p = 0.05) (Di Rienzo et al., 2011).

Table
1. Descriptions
of treatments applied for seed germination experiments.
Panicle characterization
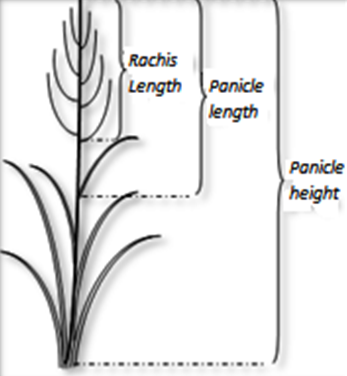
After the harvest of seeds in May of the years 2017 and 2019, the panicles evaluated for seed dehiscence previously mentioned were cut and transferred to the laboratory where the following measurements were made: Panicle height (h panicle): length in centimeters from the base of the panicle to the apex. Panicle length (l panicle): measurement in centimeters from the apex to the node of the flag leaf. Rachis Length (l rachis): distance in centimeters from the upper end to the first branch of the panicle (Figure 2). The panicles were weighed (w weight) from the apex to the node of the flag leaf, discarding the leaves and brought to an oven at 60 °C for 72 hours. Then, they were weighed (d weight) again to obtain the dry matter (%dm) of it. The normality of the data was verified by means of the Shapiro-Wilks test and then a two-ways ANOVA was performed to evaluate the effect year, genotype and its interaction for each variable (p = 0.05) (Di Rienzo et al., 2011). To visualize the joint effect of year and cultivar on all the panicle attributes, a multivariate principal component analysis (PCA) was used, which also allowed identifying those variables that explained most of the total variability contained in the data and exploring the correlations among them. Finally, a PCA per year was applied with individual plants, in order to visualize the variation structure and compare the positions of cultivars in the biplots for data from each year vs. joint data (Di Rienzo et al., 2011).
Figure 2.
Morphological panicle related traits evaluated in thos research.
Results
Seed dehiscence was lower in first weeks than in the subsequent ones in both cultivars but, as shown in Table 2, significant differences between cv, Kapivera and Bambatsi were found in the two years of evaluation (χ²= 18.11; p < 0.05 in 2017 and χ² = 30.11; p < 0.05 in 2019). However, in Figure 3 the accumulated percentage of fallen seeds to each cultivar was plotted, observing a similar behavior between them in both years until week 4, from which differences in dehiscence began to be seen until week 7 in 2017 and 6 in 2019. The Bambatsi cultivar produced a greater amount of total seeds (Table 2), so in absolute numbers throughout the two years of study it showed a more notable dehiscence. However, when the values were calculated in proportion, it was observed that between week 5 and 7 in 2017, and between 5 and 6 in 2019, the accumulated percentage of fallen seeds was higher in Kapivera (Figure 3). The peak of fall for the data set for the years 2017 and 2019 occurred on the same date for both cultivars (first days of May) although this means, as previously presented, periods of different weeks.

Table 2. Dinamic of seed dehiscence by week, individually accounted, for both cultivars at both years of evaluation.
Figure 3. Dynamics
of accumulated percentage of fallen seeds per cultivar.


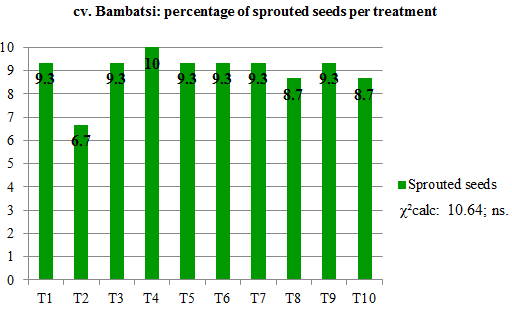
With respect to germination, no significant differences were found between cultivars for the different treatments (Figure 4) or between treatments within the cultivar (Figure 5), considering all assayed seeds in both cases. The percentage of germinatios was higher than 80 % for both cultivars.
Figure 4. Percentage
of germinated seeds per cultivar.
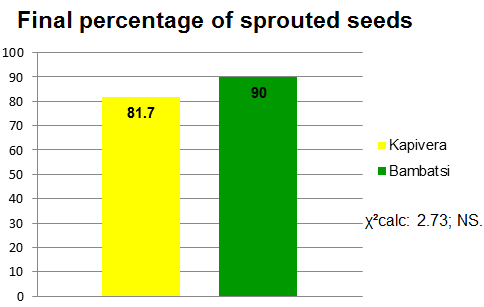
Figure 5. Percentage
contribution of each treatment to total germination per cultivar.

Respecting panicle related traits, for years 2017 and 2019, all the variables analyzed using the Shapiro-Wilks test were normally distributed, with the exception of dry weight, which had to be transformed by the logarithmic function. There were no significant differences between cultivars or between years for the panicle height, wet weight and dry weight variables, while for panicle length, rachis length and dry matter percentage, significant differences were found between cultivars and between years (Table 3). The cultivar interaction per year was not significant for any variable. For the variables in which the two-ways ANOVA detected significant differences, these were always of greater magnitude for the cultivar classification criterion than for the year one, cv. Kapivera presented the smallest values of panicle length and rachis and the highest percentage values of dry matter. On the other hand, in 2019 the values of the three variables were significantly higher than in 2017.

Table 3. Mean average values and standard deviation of the analyzed
panicle related traits (p = 0.05).
In the Principal Component Analysis (PCA) of all panicle data as a whole (Figure 6), in which cultivars and years were combined as sources of variation resulting in 4 points to visualize (Kapivera 2017, Kapivera 2019, Bambatsi 2017 and Bambatsi 2019), the first two Principal Components (PC1 and PC2) explained 99.6% of the total variability observed (68.7% for PC1 and 30.9% for PC2). Positive, high and significant correlations were found between panicle length, rachis length and wet weight, being these variables together with dry weight those that have contributions greater than 0.45 to the composition of PC1. Regarding PC2, the greatest contributions to its composition were made, positively, by percentage of dry matter and panicle height (Table 4). In agreement with these observations, in Figure 6 it is observed that cv. Bambatsi (2019) is more to the right of the graph, being more extreme in 2019 than in 2017 because, as previously mentioned, this cultivar and this year had the highest values for panicle and rachis length, variables closely associated with PC1 and that showed significant differences between years and between cultivars. On the other hand, Kapivera (2017) is more to the left of PC1, since its panicle and rachis length values were significantly lower. Kapivera had in general a higher percentage of dry matter (a variable highly associated with PC2) than Bambatsi, differences that were indeed more noticeable in 2019.
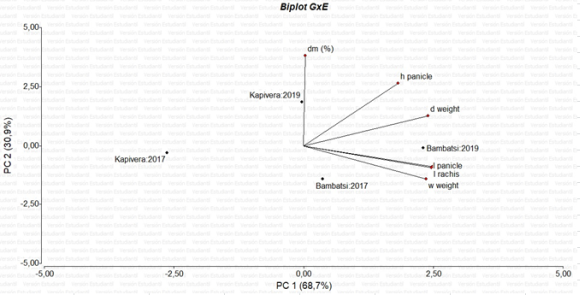
Figure 6. Biplot of Principal Component Analysis appled to data as a whole.
Table 4. Eigenvalues (Lambda) of the joint Principal Component Analysis for the panicle related traits in both cultivars assayed.

With regard to the Principal Component Analysis per year and also including all panicle data, in 2017 the first PC explained 52.5% of the total variability observed; and PC2, 22.9% (in total, 75.4%). The variables rachis length and panicle length contributed positively to both PC1 and PC2, while panicle height, dry weight and wet weight contributed positively to PC1 and negative to PC2. Percentage of dry matter, on the other hand, contributed negatively to both PC1 and PC2 (Table 5).

Table 5. Correlations of the first two Principal Components and the original variables in both PCA analysis.
Concomitantly, in the space defined by both PCs (Figure 7) it is observed that the plants of cv. Bambatsi tend to be at the top of the graph, since they presented panicle length and rachis values significantly higher than those of cv. Kapivera (whose plants tend to appear at the bottom of the graph). On the other hand, high and positive correlations were found between the variables dry weight and wet weight (correlation coefficient: 0.96), and rachis length and panicle length (correlation coefficient: 0.86) in 2017. For the year 2019, the behavior of the variables was similar to that of 2017, although the percentage of total variation explained by both PCs is slightly higher (76.7%) due to an increase in the proportion attributed to PC2 (24.2%, maintaining PC1 52.5%). In fact, this year the negative contribution of dry matter percentage to PC2 is lower than in 2017 (Table 5), achieving a greater differentiation (Figure 8) among the plants of cv. Bambatsi (which tend to be in the upper right sector of the graph) of those of cv. Kapivera, that tend to be in the lower left corner of the graph. The correlation structure in 2019 is maintained with respect to 2017 (rachis length with panicle length showed a correlation coefficient of 0.82; while dry weight and wet weight presented a correlation coefficient: 0.93), showing slight modifications that may explain the changes observed between both biplots. It should be noted that for both years, adding a third main component, the total variability explained in the data increases to 91%, continuing to be less, however, than that explained with only two PCs in the joint analysis.

Figure 7. Biplot of Principal Component Analysis applied to individual plants evaluated on 2017. Kapivera plants are marked in blue and Bambatsi plants are marked in yellow.

Figure 8. Biplot of Principal Component Analysis applied to individual plants evaluated on 2019. Kapivera plants are marked in blue and Bambatsi plants are marked in yellow.
Discussion
In Argentina, the northern Pampeana region is characterized by its high temperatures, concentrated rains and high evapotranspiration in the summer. It also shows low temperatures and lack of rain in the winter. The presence of poorly developed soils, with low organic matter content, without structure, with low moisture retention and in many environments, saline and/or waterlogged soils, are important factors to consider when choosing a forage species. Due to these characteristics, advancing in the search for species having persistence, high productivity, resistance to grazing and forage quality is necessary (Pérez, 2005).
Numerous studies on phenotypic characters have been reported in different forage grass species of great agronomic interest, evidencing the importance of selecting those species that are useful in intensive systems (rearing/wintering) with extreme conditions of abiotic stress highlighting their implications for improvement plant genetics and seed production. The amount of product that a farmer obtains for each unit of surface depends on multiple natural and human factors. Forage production does not necessarily predict the secondary production of meat, milk or wool (Meissner, 1997; Smith et al., 1997). In this sense, seed dehiscence has always been a major component of yield loss in many cereal crops worldwide.
Regarding caryopsis dehiscence, it is widely variable among the Poaceae family ranging for complete retention in crops as maize to dispersed drop along time in weeds as Echinochloa spp. (Balanzà et al, 2016). In this work a different dynamics of seed drop could be observed for each cultivar in both years under study, coinciding the peak of fall for the set of data during the first days of May (although this means different weeks) without evidencing influence of external climatic factors. These values regarding a unique drop peak differ from that reported by Dreher et al. (2008), who evaluated the dehiscence on Panicum coloratum var. makarikariense from different areas of Argentina (Córdoba and Corrientes) finding respectively two seed drop peaks influenced by intense winds and rain storms. In this case, the authors have suggested that the seeds must have been mature for dehiscence to be affected by environmental factors. In correspondance to Burson et al. (1983), this research also reports that the first seeds began to come off the mother plant 15 days after flowering began. The results obtained in the germination analysis did not demonstrate effects that respond to a maturing asynchrony, as expected from the differences in drop period among collected seeds, present in the cultivars, even when the seeds were evaluated under different scarification methods and growth conditions. These results agree with what was reported by Maina (2012). The aforementioned results were unexpected if considering that mature and dormant seeds were evaluated, according to Tischler y Young (1987) for Panicum coloratum. For this reason, it is proposed that the existing differences in the dynamics of seed drop did not affect its subsequent germination, observing high percentages of germination in both cultivars. In accordance with what was reported by Tischler y Ocumpaugn (2004), the high percentages of germination obtained suggest that the seeds are mature at the time of detaching from the mother plant, regardless of when this happens. This information also agrees with what was previously reported by Dreher et al. (2008) referring to the maturity of the seeds prior to their dehiscence.
The evaluation of the panicle related traits indicated that the differences between cultivars with respect to the characters "panicle length" and "rachis length" were related to the architecture of the vegetation, since the cultivar Bambatsi presented taller plants and more erect than the Kapivera cultivar (Dreher et al., 2008). It would be interesting to explain in another study, if the greater number of produced seeds in the Bambatsi cultivar was due to the greater size of panicle, expressed in the high values of "panicle length" obtained. As Balanzà et al. (2016) proposed, a shortest panicle would allow a more uniform fruit maturity and consequently, a more concentrated caryopsis drop. However, it is noticeable that the percentage of germination was high (greater than 80 %), which is an interesting attribute for seed production. The differences found between cultivars for the “dm (%)” character have strengthened the visual appreciation that the Kapivera cultivar develops a fundamentally creeping growth habit, with higher biomass production covering the groove. Likewise, the highest values of the variables expressed for the year 2019 were exclusively due to the ontogeny of the plants.
Through a joint principal components analysis, it was possible to explain the entire variation of the data with the first two principal components due to the high correlation between variables. In the biplot graph, the PC1 associated with the year of study could be defined, and the PC2 directly associated with the cultivar. It could be said that this last main component is the one that differentiates the years 2017 and 2019. Corroborating the results obtained in the univariate analysis and in the interaction analysis, it is important to highlight the location of the representative points of the year 2019 on the PC1 to the right of the graph. The PC2 allowed discriminating both cultivars in different years, probably due to the significant genotype x environment interaction exhibited by the variables that made the greatest contribution to this PC.
On the other hand, the PCA for each year maintains the importance of each characteristic evaluated with respect to the joint analysis due to the lack of significant GxE interaction observed in general (due to general variability). In addition, it highlights the absence of a defined grouping of the plants with respect to the cultivar to which they correspond, which is more noticeable in 2017 than in 2019, possibly because the early evaluation was less representative of the differences between cultivars than a later one (Senanayake et al., 1994).
It is logical to find these results assuming that the Kapivera cultivar has been developed from a selection within the Bambatsi cultivar. Expected correlations between a certain group of variables can also be visualized, which also, according to the length of the vector, seem to have similar importance when discriminating cultivars. As suggested by Balanzà et al. (2016) in Arabidopsis, hemce there is probably a conception based on the fact that the smaller size of panicle leads to a reduction in the maturational asynchrony of the seeds in Panicum coloratum var. makarikariense, naturally resulting in a decrease in its dehiscence. This is supported by the lower values found for the variables related to panicle ("l panicle", "h panicle" and "l rachis") in the Kapivera cultivar, resulting in a reduction in total produced and fallen seeds compared to the Bambatsi cultivar. In addition to these results, it is probable that better climatic conditions have allowed to express their differences in the phenotype of the plants (Schlichting, 1986).
Due to the limited progress in the knowledge of the behavior of the main phenotypic characters associated with the yield of Panicum coloratum var. makarikariense, it is essential to provide information about them in order to offer new tools that assist the genetic improvement of crops in the search for an increase in grain yield and in the highest quality of fruits obtained. However, it is necessary to mention that seed production in yield was not evaluated in thos work, neither its relations with panicle related traits. Seed production was assessed in terms of morphological and phenological traits, such as caryopsis dehiscence, percentage of germination, and panicle attibutes.
Conclusions
Among the two characterized cultivars of Panicum coloratum var. makarikariense (cv. Kapivera and cv. Bambatsi) in the Argentinean Pampeana Region, with a temperated climate, no differences were found for the germination behavior and the biomass production traits panicle height, wet weight and dry weight. Instead, differences in the dynamics of seed dehiscence, panicle length, rachis length and dry matter percentage were detected between cultivars and years of evaluation but the genotype x year interaction was non-signifficant.
Acknowledgments
The plant materials of Panicum coloratum were contributed by the National Institute of Agricultural Technology (INTA), Rafaela Experimental Station. The experimental lot on which the materials were transplanted was transferred by the Department of Forages (Faculty of Agricultural Sciences).
Funding statement
This work was financed by the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) grant P-UE: 22920160100043CO (IICAR), and by the National University of Rosario, Projects of Technological Linking and Productive Development “Inclusive Linking” 2015. GR Pratta and SA Felitti are research career members of the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Conflict of interest
The authors declare that they have no conflict of interest.
ARMANDO, L. V.; CARRERA, A. D. y TOMAS, M. A. (2013). Collection and morphological characterization of Panicum coloratum L. in Argentina. Genetic Resources and Crop Evolution, 60(5), 1737–1747. https://doi.org/10.1007/s10722-013-9982-3
BALANZÀ, V.; ROIG-VILLANOVA, I.; DI MARZO, M.; MASIERO, S., y COLOMBO, L. (2016). Seed abscission and fruit dehiscence required for seed dispersal rely on similar genetic networks. Development, 143(18), 3372-3381.
BURSON, B. L.; CORREA, J. y POTTS, H. C. (1983). Anatomical Basis for Seed Shattering in Kleingrass and Guineagrass1. Crop Science, 23(4), 747. https://doi.org/10.2135/cropsci1983.0011183x002300040035x
CUBERO, J. I. (2003). Introducción a la mejora genética vegetal (2013th ed.). Mundiprensa.
DI RIENZO J. A.; CASANOVES F.; BALZARINI M. G.; GONZALEZ L. y TABLADA M., R. C. W. (2011). InfoStat. Grupo InfoStat. FCA, Universidad Nacional de Córdoba, Argentina.
DREHER, N.; PISANI, M.; TOMÁS, M. A. y BERONE, G. (2008). Producción y dinámica de la dehiscencia de semillas en poblaciones de Panicum coloratum var. makarikariensis. Revista Argentina de Producción Animal, 28, 349–543.
FOSSATI, J. L. y BRUNO, O. A. (1982). Comportamiento de especies forrajeras de clima templado y subtropical en el centro oeste de la provincia de Santa Fe. In INTA Publicación técnica (Vol. 21, p. 43).
HACKER, J. B. y LOCH, D. S. (1997). Tropical forage seed production: producer’s views and research opportunities. XVIII International Grassland Congress, Winnipeg.
HAYES, P.; STUCKER, R. y WANDREY, G. (1989). The domestication of American wildrice. Economic Botany, 43(2), 203–214.
INDEC. (2002). Superficies implantada en las EAP con y sin límites definidos, por grupo de cultivos, según período de ocupación y provincia. Retrieved from https://www.indec.gob.ar/indec/web/Nivel4-Tema-3-8-87
KENNARD, W. C.; PHILLIPS, R. L. y PORTER, R. A. (2002). Genetic dissection of seed shattering , agronomic , and color traits in American wildrice (Zizania palustris var . interior L .) with a comparative map. Theoretical and Applied Genetics, 105(6–7), 1075–1086. https://doi.org/10.1007/s00122-002-0988-z
LOCH D. S. y FERGUSON J. E. (1999). Forage Seed Production. Volume 2: Tropical and Subtropical Species. In Tropical and subtropical forage seed production: An overview (p. 10). CAB International.
MAINA, M. (2012). Variabilidad en caracteres relacionados a la producción de semilla en poblaciones de Panicum coloratum var. makarikariense. Universidad Nacional de Rosario.
MEISSNER, H. H. (1997). Recent research on forage utilization by ruminant livestock in South Africa. Animal Feed Science and Technology, 69(1–3), 103–119. https://doi.org/10.1016/S0377-8401(97)81626-4
PATTERSON, S. E. (2001). Cutting loose. Abscission and dehiscence in Arabidopsis. Plant Physiology, 126(2), 494–500. https://doi.org/10.1104/pp.126.2.494
PÉREZ, H. E. (2005). Características de las especies forrajeras adaptadas a las condiciones del NO del país. In FORRAJES 2005 (pp. 1–6).
PETRUZZI, H.; STRITZLER, N.; ADEMA, E. O.; FERRI, C. M. y PAGELLA, J. H. (2003). Mijo perenne - Panicum coloratum. In EEA Anguil INTA (pp. 1–28).
SCHLICHTING, C. D. (1986). The evolution of phenotypic plasticity in plants. Annual Review of Ecology, Evolution, and Systematics, 17(143), 667–693.
SENANAYAKE, N.; LANKA, S. y NAYLOR, R. E. L. (1994). Variation in development of contrasting rice cultivars. The Journal of Agricultural Science, 123(April 2016), 35–39. https://doi.org/10.1017/S0021859600067745
SMITH, K. F.; REED, K. F. M. y FOOT, J. Z. (1997). An assessment of the relative importance of specific traits for the genetic improvement of nutritive value in dairy pasture. Grass and Forage Science, 52(2), 167–175. https://doi.org/10.1111/j.1365-2494.1997.tb02347.x
SPENCE, J.; VERCHER, Y.; GATES, P. y HARRIS, N. (1996). “Pod shatter” in Arabidopsis thaliana, Brassica napus and B. juncea. Journal of Microscopy, 181(2), 195–203. https://doi.org/10.1046/j.1365-2818.1996.111391.x
TISCHLER, C. R. y YOUNG, B. A. (1987). Development and Characteristics of a Kleingrass Population with Reduced Post‐Harvest Seed Dormancy. Crop Science, 27(6), 1238–1241. https://doi.org/10.2135/cropsci1987.0011183x002700060030x
TISCHLER, C. R. y OCUMPAUGH, W. R. (2004). Kleingrass, Blue Panic, and Vine Mesquite. Warm-Season (C4) Grasses, (45), 623–649.
WILSON, R. G. (1963). Bambatsi grass for Downs and Brigalow. Queensland Agricultural Journal, 89, 118–119.